

社会問題と向き合う人のクラウドファンディング「GoodMorning」にて実施されたプロジェクトです。
11/24:NEXT GOALについて追記しました!
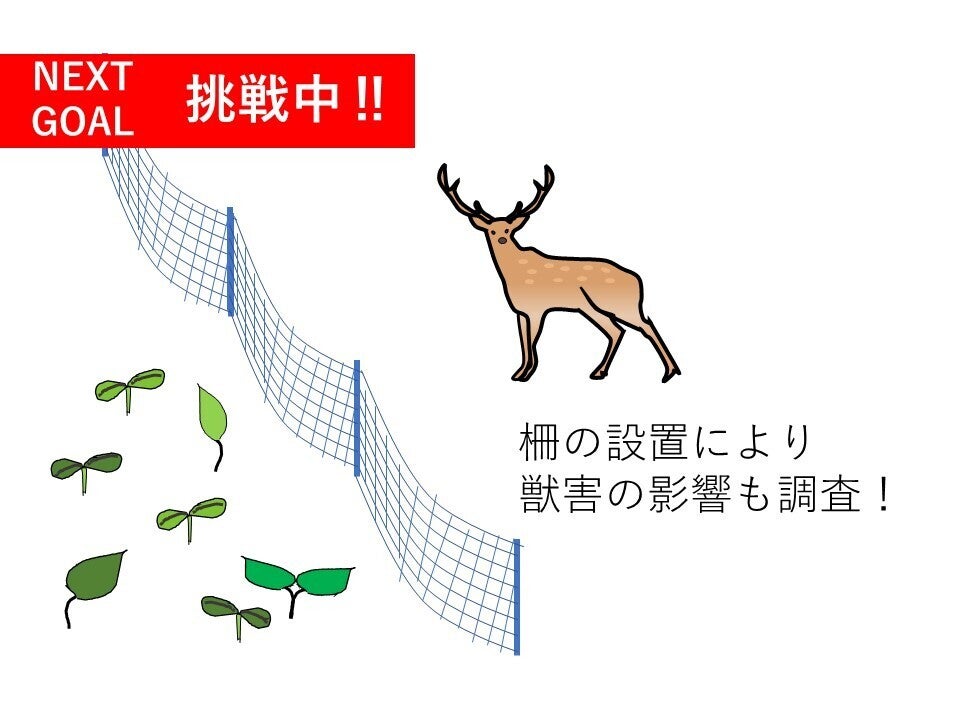
お陰さまで当初の目標金額を達成することができました!心よりお礼申し上げます!
そこで、ネクストゴールとして獣害対策用の柵設置費用を含めた総額150万円を設定することに致しました。ご存じの方もいらっしゃると思いますが、日本の森林では近年、シカやイノシシによる被害が急増しています。動物が増えることは一見すると良いことに見えますが、時には森林にとって悪い影響を与えると考えられています。シカの場合、採食行動によって若い木を食べつくし森林の更新を阻害したり、スギ・ヒノキの樹皮を食べ傷つけてしまったりという問題が発生しています。
本研究で獣害対策用の柵を設置する場所と、設置しない場所を設定することで、紀南地域における獣害が森林の天然更新へ及ぼす影響、つまり柵設置の必要性を調査したいと考えています。
ご支援どうぞよろしくお願いします!

はじめまして!今回プロジェクトを立ち上げた井口光(いぐち ひかる)と申します。

私は小さい頃から山村巡りが大好きで、出身地の東京にある奥多摩をよく歩き回っていました。山登りではなく山村巡りが好きなのは、自分の育った住宅街とは全く異なる環境、そして長い歴史を重ねてきた集落の生活がとても魅力的だったからです。
集落の周りにある森林にも興味を持ち始め、大学では森林科学を専攻しました。入学後、学科の実習で訪問した熊野の集落風景を見て、「ここで暮らしてみたい!」と一目惚れをし、和歌山県古座川町にある北海道大学の研究林で大学院生としての研究活動を始めることを決意しました。
 研究林内での調査風景
研究林内での調査風景
しかし、研究を進めるにあたり自分の力ではどうにもできない資金面での課題に直面し、皆様にご協力いただきたくクラウドファンディングを行うこととしました。
一人の学生の研究ではありますが、その結果は必ず日本全体の森林管理にとって、また皆さまにとっても価値のあるものになると確信しております。なぜ重要なのか?なぜ資金が必要なのか?皆様に納得して頂くため、本プロジェクトを始めるに至った経緯をご紹介したいと思います。

まず、私の所属する北海道大学和歌山研究林について簡単に紹介したいと思います。
和歌山県古座川町?北海道大学なのに和歌山研究林?ほとんどの方はこう思ったのではないでしょうか?
古座川町は本州最南端の町・串本町のひとつ内陸側に位置する人口約2,700人の町です。
町の名前にもなっている古座川は環境省が発表した平成の名水100選に選ばれるほどの透明度を誇り、上流域は貴重な生物(オオサンショウウオなど)の住処になっています。さらに古座川流域は「日本の秘境100選」にも選ばれ、風光明媚な光景が広がっています。
また、近年では町内に自生する標本木などを元に「クマノザクラ」という新種のサクラが発見されたことで話題にもなりました。
 古座川町の位置と町内の代表的な風景
古座川町の位置と町内の代表的な風景
 特別天然記念物オオサンショウウオ(学術目的での特別な許可を得て観察しています)
特別天然記念物オオサンショウウオ(学術目的での特別な許可を得て観察しています)
その古座川町の最も奥まった場所に、平井(ひらい)という80人ほどが暮らす小さな集落があります。
平井集落は鎌倉時代の戦乱から逃れた人々より始まる約800年の歴史がありますが、現在は古座川支流にあたる平井川の清流に育まれたユズの栽培と加工品の生産で名が知られています。
 平井集落
平井集落
北海道大学 和歌山研究林 はこの平井集落に、北海道では見ることのできないスギ・ヒノキの人工林や暖温帯特有の森林(自然状態ではシイ類・カシ類など常緑の広葉樹が生い茂る森林)に関する教育や研究を目的として、1925 (大正15) 年に創立されました。
 平井集落にある北海道大学 和歌山研究林の庁舎。1927(昭和2)年建設。登録有形文化財。
平井集落にある北海道大学 和歌山研究林の庁舎。1927(昭和2)年建設。登録有形文化財。
事務所のほか、利用者のための実験や宿泊スペースとしても機能している
 和歌山研究林の森林遠景。手前から1つ目と2つ目の稜線から広がる山域がほぼ研究林敷地。
和歌山研究林の森林遠景。手前から1つ目と2つ目の稜線から広がる山域がほぼ研究林敷地。
この北海道大学和歌山研究林に来てみると、里山周辺の奥地の人工林は管理がままならぬ事態へと陥っていることを目の当たりにしました。若者が都市部へと流出して、管理する人手が足りないからです。
調べてみると、全国各地の里山でも奥地の人工林は管理が放棄され、同じような問題を抱えていました。さらに本州では北海道よりも地形が急峻であるということも相まって、事態はより一層深刻な状況です。
授業で話は聞いていましたが、まさかここまで厳しいとは思ってもみませんでした。
この奥地の放棄された人工林を救うために何かできないだろうかと考えた結果、私たち(私と和歌山研究林職員)は人工林に隣接する天然林から常緑広葉樹の侵入を誘導し、放棄された奥地の人工林を効率よく天然林へと戻すことで、「低コストで管理できる森づくり技術」の開発を目指すプロジェクトを立ち上げました!
 和歌山研究林内の90年生スギ人工林。
和歌山研究林内の90年生スギ人工林。
林道沿いの平地などに広がる一部の場所はこのように管理が行き届いているが…

では、なぜ管理がままならない奥地人工林が増えてしまったのでしょうか?
原因は様々ですが、大きなものとして日本の林業をとりまく環境の変化があげられます。
 戦後日本の人工林の取り扱いの歴史的背景
戦後日本の人工林の取り扱いの歴史的背景
戦後の日本は、軍需物資として木材資源が切り出され荒廃した国土の復興と、高度経済成長に伴う木材需要の増加に応えるべく、山という山にスギやヒノキの苗木が植えられ、驚くほど山奥にまで人工林が作られていきました。しかし、その人工林の成長を待たずして、1960年代に安く大量に入手できる外国産材の輸入が自由化されたことをはじめとする様々な要因が重なり、国際的な競争力に乏しかった日本の林業はみるみる衰退してしまいました。
その結果、人々のスギ・ヒノキ人工林への関心が薄れ、特に管理コストがかかる奥地人工林は管理がままならぬ事態へと陥ってしまったのです。
通常、日本の針葉樹人工林は、間伐や枝打ちなど定期的に管理されることを前提に収穫目標よりも多めの本数が植栽されます。
そのため、前提とされていた管理が行き届かなくなると、林内は過密状態となり荒廃してしまいます。
 和歌山研究林内の奥地に広がる、管理がままならない状態のヒノキ人工林。
和歌山研究林内の奥地に広がる、管理がままならない状態のヒノキ人工林。
林内は暗く林床植生が乏しい。
一方で、人工林とは異なり、天然林では間伐や枝打ちなどの作業コストが不要なため、管理コストが抑えられます。
そこで、この天然林に近い形へと人工林を誘導することで、管理コストを抑えながら森林の状態を改善しようというのが、本プロジェクトの目的です。

また、天然林に近い形へと誘導することで得られるメリットは、管理コストの削減だけではありません。
現在の日本では、木材生産のみならず、生物多様性保全や河川に流れ込む水量をコントロールして洪水を緩和する水源涵養の機能など、多面的な機能を高める目的で森づくりが進められています。針葉樹人工林に比べて天然林にみられるような多くの種が存在する森は、この多面的機能が高いと言われています。
人と森林がより長く共存できる関係を築くために、人工林だけではなく、多くの樹種が生存する天然林に近い形へと戻す森づくりが必要とされているのです。

国土の2/3を森林が占めている日本で、森林の健全さは国土の健全さと言っても過言ではありません。
そのため、森林の状態は下流の都市に住む人々にも無関係ではないのです。
例えば、休日にレクリエーションに行った山が荒廃していたら、自然の恵みを感じませんよね?健全な森の方がレクリエーション機能が高いのです。
また荒廃した人工林では、しばしば土壌の流出が問題になります。この問題は河川の水質や、災害の原因にもなり直接下流の人々に影響を及ぼします(洪水の制御機能)。
この国に暮らす人にとって「最も身近で最も大きな隣人」とも呼べる森林の健康を維持することは、将来に渡って良好な“ご近所付き合い”を続けていくために必要不可欠なのです。そして本プロジェクトが成功し、広葉樹林化への手掛りを示すことが出来れば、全国の荒廃した森林の状態は改善され、より良い森林との隣人関係を築くことが出来るのです。

では実際にどのようにプロジェクト(研究)を進めるのかご紹介したいと思います。
私たちの所属する和歌山研究林には天然林と人工林があります。
これら森林を利用して、スギ・ヒノキ人工林に隣接する天然林から常緑広葉樹の侵入を誘導し、里山奥地の放棄された人工林を効率よく天然林へと戻す森づくり技術を開発したいと考えています。
 和歌山研究林の最奥部にある、スギ人工林(左)と常緑広葉樹(照葉樹)の天然林(右)の境目。
和歌山研究林の最奥部にある、スギ人工林(左)と常緑広葉樹(照葉樹)の天然林(右)の境目。
スギ人工林側は手入れがあまり行き届いておらず、細く枯れ枝の多い木が多く残っている。
 境目を上空から撮影。右上が常緑広葉樹林、左下が人工林
境目を上空から撮影。右上が常緑広葉樹林、左下が人工林
さて、ここでまた問題が生じます。どのように広葉樹を誘導すれば良いのでしょうか?
ぱっと思いつくのは人が植えることでしょう。
しかしこの方法では、苗木を山の上まで持っていき、穴を掘って植栽し、下刈りなどの作業を続けながら、時間をかけて手厚い世話を続けなければなりせん。ましてや人手の少ない里山奥地において、このような作業を続けていくことは、支払うコストがことさらに大きくなってしまいます。そもそも管理コストを下げることが目的なので、これでは本末転倒です。
 天然更新により回復しつつある皆伐跡地
天然更新により回復しつつある皆伐跡地
そこで私たちが注目したのは、天然更新です。
天然更新とは簡単にいうと、‶植えても種を播いてもいないのに、勝手に生えてくる木を使って森を作る”ことです。木は本来、鳥や風、動物の力、重力を借りて種子を拡散することで、世代交代する力を持っており、そのお陰で地球上に森林が維持されてきたという背景があります。この元から木が持っている世代交代する能力を使って、森を作ろうというのが天然更新です。
私の研究ではこの天然更新を利用し、広葉樹を誘導しようとしています。
 鬱蒼とした林内に倒木が発生するなどしてできた、小さな「日なた」の部分。
鬱蒼とした林内に倒木が発生するなどしてできた、小さな「日なた」の部分。
森林生態学の世界では「ギャップ」と呼ばれ、日本のような温暖多雨な気候においては、
このギャップにさす陽の光によって次世代の若い森林が育っていく。
そしてこの天然更新には光が重要になります。
十分な光がなければ、植物は成長することが出来ないからです。
そこで「光」を十分に地面に届くように間伐をすることで、広葉樹が天然更新しやすい環境を整えることにしました。

通常、人工林ではスギ・ヒノキの枝葉が空を覆っているため林内は暗くなっています。
この暗い状況は、侵入した広葉樹が種子から発芽したり成長したりするには、非常に過酷な環境です。そのため針葉樹を部分的に間伐し、空を開けて明るくすることで、「広葉樹が発芽・成長しやすい環境」を作り出せると考えられます。
しかし、切れば切るほど良いというわけではありません。
切りすぎると、地面が乾燥したり、降雨による表土の流出が起こったりとマイナスな面が顕著になってしまいます。
これらの兼ね合いを見極め、最適な間伐強度を見つけだすことが、この研究のポイントになります。最適な間伐強度が分かれば、自然の力を借りて、効率的に、低コストで、管理できる森林へと針葉樹人工林を転換できる方法を示せるのです。そしてこの方法を示すことが出来れば、全国の荒廃した針葉樹人工林を改善するための技術開発を一歩進めることが出来るのです!
 本プロジェクトの実験デザイン。様々な強度(幅)で間伐を行うことで、常緑広葉樹が侵入しやすい最適な強度を見つけ出すことを目的にしている。
本プロジェクトの実験デザイン。様々な強度(幅)で間伐を行うことで、常緑広葉樹が侵入しやすい最適な強度を見つけ出すことを目的にしている。

しかし、大きな課題が残っています。
「和歌山研究林には実験を行うために必要な林道・作業道などの路網が十分に整っていない!」
ということです。
一般に、古座川流域のような急傾斜地の多い山林において、伐採などの林内作業を行うためには最低でも1ヘクタールの山林面積あたり、50mの作業道(林道よりも簡易な構造で、積載量2tのダンプが通行できる道路)が必要とされます。しかし現在の和歌山研究林においては、1ヘクタールの山林面積あたり、20mの道路網しか整備されていません。しかも、既設の道路網の大半は林内を縦断・横断している谷底の河川沿いに集中しています。
 一部の沢沿いにはこのような林道が敷かれているが…
一部の沢沿いにはこのような林道が敷かれているが…
 研究のための実験サイトの検討のため、路網が整備されていない約40度の傾斜地を歩く筆者。
研究のための実験サイトの検討のため、路網が整備されていない約40度の傾斜地を歩く筆者。
この直後に浮石を踏んで滑り落ちてしまい、腕や足に擦り傷が…

加えて、田辺市本宮町の川湯温泉などで壊滅的な被害を出した2018年8月23日の台風20号の影響により、和歌山研究林の既存の道路網の多くが土砂災害で破壊されてしまいました。
幸い、破壊された林道などは少しずつ修復を進めている状況ですが、土木資材の在庫品の一部が道路と共に流失してしまったこともあり、和歌山研究林だけの力では新たな作業道の作設は難しい状態です。
 台風20号による林道の大規模な崩壊。土石流により林道のほかに物置や橋梁などが押し流されてしまっている(2018年8月撮影・現在は一部復旧済み)
台風20号による林道の大規模な崩壊。土石流により林道のほかに物置や橋梁などが押し流されてしまっている(2018年8月撮影・現在は一部復旧済み)
もし林道が整備できなかったとしたら、研究が難航するだけでなく、伐採した木材を放置することにもなります。
スギ・ヒノキなどの針葉樹人工林は、植えられた苗を育てて伐採し、最終的に木材として利用するための森林です。しかし、現状は伐採された木を運び出す路網がないため、間伐された原木を山の中に残したままになってしまいます。
これでは丹念に苗を植えた先人の方々に、まったく申し訳が立ちません。
 急傾斜地における先人たちの丹念な苗の植付け作業(1970年代・和歌山研究林所蔵)
急傾斜地における先人たちの丹念な苗の植付け作業(1970年代・和歌山研究林所蔵)

研究が終了した後も、新たに開設した林道や搬出された木材は環境教育を行っている研究林では価値を発揮します。
例えば、研究林には木材を加工する設備があるため、森の木を切り、林道から搬出できるようになると、製品として加工・販売するまでの一連の作業をすることが可能になります。
つまり、「川上で行われる木材生産から、川下で行われる商品加工・販売までの流れ」を一度に見ることができます。
加えて、林道が整備されることで車両を使った案内が可能となり、森林と関わる機会が少なくなった子供たちを含めより多くの方々に「研究林内の様々な取り組みを体験する教育プログラム」も提供できるようになります。
また今日的な意義としては、森林の中は「3密」を避けた状況を作りやすく、withコロナの時代に適した教育プログラムが実施できるという利点も兼ね備えています。
 森林の中に路網が新設されたら、木材加工や教育サービスの提供が可能になる
森林の中に路網が新設されたら、木材加工や教育サービスの提供が可能になる

本プロジェクトでは、森づくり技術開発に必要な作業道新設のための土木資材の購入費90万円を、私たちだけで用意することができず、皆様のご協力を必要としています。
作業道の土木資材が準備できれば、その他必要な重機や道具は和歌山研究林に揃っています。
資材さえあれば!あとは自分たちで進められる状態なのです。
「里山奥地の人工林問題の解決に向けた森林づくり技を開発する」ために、さらに「先人の植えた森林資源を大切に利用する」ために、和歌山研究林の路網新設は不可欠なのです。
ご支援のほど何卒よろしくお願いいたします!

本プロジェクトを立ち上げるにあたって、和歌山研究林とご縁のある多くの方々から応援メッセージをいただくことができました。皆様、本当にありがとうございます!
★西前啓市 様 (古座川町長) より

古座川町長の西前啓市でございます。
本町は、面積の約96%が森林であり、林業が盛んな時代には、林業に携わる者も大勢おりましたが、現在では、林業従事者も少なくなり、手入れの行き届いていない人工林が増えていることも事実です。
森林を再生させるには、50年100年という長い期間を要し、林業従事者の人材確保が必要となります。
本町におきましても森林経営管理法に基づき森林整備を進めているところでございますが、今回のプロジェクトである、「低コストで管理できる森づくり技術」の開発が出来れば、本町の抱える荒廃した奥地人工林の整備に大変有益であろうと期待しております。
ぜひ実現に向けて、頑張って頂きたいと思います。
★きのくに子どもの村中学校 動植物研究所の皆様 より

きのくに子どもの村中学校の動植物研究所です。
毎年クラスで北海道大学和歌山研究林のみなさんにお世話になり、自然林の観察や小動物の捕獲・観察、間伐体験などをさせていただいてきました。様々な手厚いサポートにとても感謝しています。ここならではの貴重な体験ができて中学生たちは目を輝かせています。今回の自然林回復のためのプロジェクトをお聞きして、とても興味をもちました。ぜひまた、いろいろ教えてください。
★ 井上節子 様 (NPO法人和歌山芸術文化支援協会(wacss) 理事長)より

「アートは人、町を元気にする」をコンセプトに2001年より様々なアート活動を続けています。
2007年より、歴史と文化を育んできた熊野の森にアーティストを招き、滞在制作や作品を通していつもの風景が違って見えたり、心の中で何かが動き出したり、森からのメッセージを体感して欲しいと願いアートプロジェクト「森のちから」を実施しています。
11回目となる「森のちからⅪ・森の中へ」は2019年10月、北海道大学和歌山研究林のスタッフの皆さんの真っ直ぐな好奇心に支えられ素晴らしいプロジェクトとなりました。
人口林から天然林へ、熊野の森の再生は未来へつながると信じています。それぞれの立場で「森に心を寄せる」今回の取り組みを応援します。
★農事組合法人 古座川ゆず平井の里の皆様 より

北大和歌山研究林さんのお隣で、特産品のユズを使った製品を作っています。
私たちの製品は、ユズの生育に適した平井集落の気候・ユズ畑のお世話や収穫をしてくださる方々・そして周辺の山々に広がる森の中より注がれた豊富な水資源に支えられています。このため、森林の健康を維持するということは、巡り巡って私たちの健康を維持することにもつながることだと日々、とても強く感じています。
山の恵みを少しずつ頂戴しながら、将来にわたって良好な「森とのご近所付き合い」を続けていくために、私たちからも応援させていただきます!
★中村誠宏 様 (和歌山研究林長・准教授) より

和歌山研究林の中村です。
井口くんの指導教員として研究デザインなど日頃から良く研究の議論をしています。私の専門は昆虫の群集生態学ですが、彼の情熱に押されて、林業にも強く関心を抱くようになりました。彼の研究から「最適な間伐強度を探る」ことで広葉樹林化するための詳細な条件を提示でき、既存の研究とは一線を画した斬新さがあると思います。
林業の研究は結果が出るまでに時間がかかります。彼の修士論文では初期の一部だけしか研究できないかもしれません。しかし、このプロジェクトで作業道が新設されれば、徒歩や自動車で実験プロットまで簡単にアクセスできるようになり、一般の方々も楽しく調査に参加できる「シティズン・サイエンス(市民参加型の科学実験)」へ展開できるのではと期待しています。

CAMPFIREさんの手数料を除いた全額を、本プロジェクトの資金として大切に活用させていただきます。いただいた資金の大半は作業道の作設に必要な土木資材や重機のレンタル代に充てるほか、リターンの送料やパッケージング等のプロジェクト運営費にも一部使用させていただきます。
 小型重機で林道の補修を行っている様子(和歌山研究林内)
小型重機で林道の補修を行っている様子(和歌山研究林内)
以下が大まかな内訳です:
・土木資材(ふとんかご・大型土のう他) ¥35万
・小型重機(3tクラスのミニバックホウ)レンタル料&燃料代 ¥30万
・プロジェクト運営費(リターン送料他) ¥11万
・手数料14% ¥14万
・総計 ・・・ ¥90万

大まかには、以下のスケジュールで進めていく予定です。
実験や道づくり、リターン制作などの様子は随時、情報発信していきます。
・2020年11月 プロジェクトページ開設、間伐実験の開始
・2021年2月~ 作業道の作設開始
・2021年4月~ 木製品などの制作・リターン発送
・2021年5月~ 野外セミナー開催 (※)時期は前後する場合があります
・2021年10月頃まで 作業道の作設完了
・2022年3月(予定)~ 研究報告書の発送 (休学などで時期が遅れる場合があります)
(※)本プロジェクトはAll-in方式で実施します。目標金額に満たない場合も、計画を実行し、リターンをお届けします。

本プロジェクト、そして和歌山研究林の特徴を最大限に活用した、以下の4種類のリターンを用意させていただきました。
(1)井口の研究成果をまとめ上げた研究報告
本プロジェクトで実施される研究の後にまとめ上げることになる研究報告をリターンに設定させていただいています。
研究の成果物という特性上、発送は最短でも2022年3月以降となりますが、休学などの諸事情により発送時期が遅れる場合があります。

(2)材料の調達から加工まで全て和歌山研究林スタッフの手で作られた木製の工芸品
和歌山研究林に生えている樹木はスギ・ヒノキ造林木のほかに180種を超える自生種がありますが、その一部は風倒などの被害が出た際に木材として切り出され、工芸品などの原料として活用されています。
この和歌山研究林産の木材を使った工芸品を、リターンのひとつとして用意させていただきました。
レーザーカッターなどのシステマチックな工具は一切使用しておらず、材料の調達から最後のひと磨きに至るまで、全て研究林スタッフの手により木材の特性を見極めながら一つ一つ丁寧に仕上げています。
 和歌山研究林スタッフによる工芸品の制作風景
和歌山研究林スタッフによる工芸品の制作風景
また先にも申し上げましたように、和歌山研究林では2018年8月に甚大な台風被害に見舞われました。その際に林道の崩落など多くの被害があったほか、多数の風倒木も発生しています。
リターンとなる木製品の一部には、この風倒木を原材料としています。
普段は古座川町の特産品として 道の駅一枚岩 Camp&Food monolith 様でのみ販売されているものですが、今回は本プロジェクト限定品としてのアレンジを加えた形で3種類提供させていただきます!
[1] コースター(2枚セット)
 (※)後ろのグラスは付属しません
(※)後ろのグラスは付属しません
[2] 天然ケヤキの樹皮を使った動植物の壁掛け

[3] スマホスタンド
 (※)後ろのスマホは付属しません
(※)後ろのスマホは付属しません
(3)和歌山研究林庁舎の設計図を元に作成した手ぬぐい
登録有形文化財である和歌山研究林の庁舎は、1927(昭和2)年に竣工されて以来、100年近くにわたり大規模な改修やリフォームなどを行うことなく現役で使用されています。
この庁舎が建設される際に描かれた本物の設計図の一部をトレスしてデジタル化し、図柄のデザインに起こした手ぬぐいをリターンのひとつに設定させていただきました。
 手ぬぐい図柄(※デザインは若干変更される場合があります)
手ぬぐい図柄(※デザインは若干変更される場合があります)
トレスされた設計図に書かれている日付はなんと、1925(大正15)年!
これは現在の和歌山研究林にあたる施設が創立された年で、当時は海沿いにある古座の町から徒歩で丸1日かけて現地へアクセスしていたそうです。
手ぬぐいという実用品を通じて、当時の雰囲気を少しでも感じ取っていただけましたら嬉しいです。
 庁舎設計図の一部。当時の学校名は「北海道帝國大学」
庁舎設計図の一部。当時の学校名は「北海道帝國大学」
(4)和歌山研究林で実施される野外セミナーへのご招待
和歌山研究林の主要なミッションは森林フィールドを使った教育・研究サービスの提供ですが、都市部から遠く離れた立地ということもあり、その具体的な内容は世間一般になかなかイメージし難い部分があります。事実、地域住民の方のなかでも和歌山研究林でどのような活動が行われていのるかあまり知られておらず、ましてや森林を所有している大学があるということ自体も、それほど広く認知されていません。
そこで!
研究林スタッフの皆様による全面的なご協力のもと、本プロジェクトの進捗状況も兼ねた、和歌山研究林のフィールドを隅々まで活用した野外セミナーをリターンとして用意させていただきました。
 野外セミナーのイメージ (過去に実際に行われた学生実習などの風景)研究紹介・林内調査の体験・急傾斜に敷かれた小型モノレール試乗・木工ワークショップ・バーベキューや窯作りからのピザ焼き…等々。
野外セミナーのイメージ (過去に実際に行われた学生実習などの風景)研究紹介・林内調査の体験・急傾斜に敷かれた小型モノレール試乗・木工ワークショップ・バーベキューや窯作りからのピザ焼き…等々。
日帰り・あるいは1泊2日(宿泊場所は研究林庁舎内)の日程で、自然の仕組みや森林環境の役割・山の恵みの利用などについて、文字通り五感をフルに使って楽しく学んでいただく機会を提供させていただきます。
山林での活動からイメージできる一般的な「山仕事」や「林業」とは一線を画す、大学が所有する森ならではの、様々な視点から森林について知ることができる体験プログラムをぜひ、満喫してください。

私たちは、「里山奥地の人工林問題」を解決するために、低コストで管理できる森づくり技術の開発と普及を目指しています。
皆様から少しでも多くの応援とご支援をいただけることによって、新たな「森づくり技術」の開発が実現化して、この人工林の問題が解決に向かうことを信じています。
精一杯がんばりますので、ご支援のほど何卒よろしくお願いいたします!

 サポートしてくださる和歌山研究林スタッフ&学生と一緒に集合写真
サポートしてくださる和歌山研究林スタッフ&学生と一緒に集合写真
★ 井口光(いぐち ひかる)

私達のもっとも身近で最も大きな隣人である森林を健康にしたい!
その思いで本プロジェクトを始めました!どうぞご協力お願いいたします!
井口光の活動歴/実績
北大農学部卒。実習で訪れた和歌山研究林の風景に一目惚れしてしまい、大学院生活を和歌山で送ることを決意。札幌では部活でアイスホッケーに打ち込みつつ、休日になると北大構内に住むキツネの一家を観察していた。その際に撮影した写真が日本政策投資銀行の北海道版パンフレット(2019年版)の表紙や北大公式カレンダーに採用された。現在は研究活動の傍ら、古座川町内の小学校で行われている放課後活動支援などを通じ、教育機関としての研究林の魅力を伝えている。趣味は筋トレ。器具がないので最近は専ら自重トレーニングをしている。好きな木はホオノキ(朴木)
 札幌キャンパス在学中に撮影していたキツネの一家。この年は子供が5匹ほど生まれ賑やかだった
札幌キャンパス在学中に撮影していたキツネの一家。この年は子供が5匹ほど生まれ賑やかだった
サポートする和歌山研究林スタッフ紹介
和歌山研究林には合計9名の技術スタッフが所属しており、今回の研究を全面的にサポートしてくださいます。このなかで特に関わりの深い5名の方々について、簡単にご紹介いたします。
★ おがわ あきふみ
…リターン製作・企画 担当
北海道出身・理学部数学科を卒業して森林の世界に入る・木材利用について研究林内で最も熱い情熱を注ぐ・好きな木はハリギリ(針桐)。
★ ちい よしたか
…間伐実験の実働・リターン製作 担当
大阪からIターンで古座川町に定住・チェンソー等を使った伐倒技術の指導者としても全国的に活躍・好きな木はケヤキ(欅)。
★ むろ たかはる
…間伐実験の実働・リターン製作・コンテンツ製作補助 担当
地元、古座川町出身・周辺地域の情報に精通し、定住希望者のアドバイザーや観光ガイドなどを歴任・特技はロックバランシング・好きな木はクロバイ(黒灰)。

★ まえだ じゅん
…リターン製作・企画 担当
地元、平井集落出身・平井在住者の中では最も若手のひとりだが和歌山研究林を最も愛し、その歴史を誰よりも広く深く知る・好きな木はキリ(桐)。
★ いとう きんや
…企画・コンテンツ制作補助 担当
千葉県出身・元SE・以前の配属先は毎年冬に-30℃未満を記録する極寒の地・関東出身だが内地の夏の暑さが苦手・好きな木はメタセコイア(曙杉)
最後までお読みいただき、ありがとうございます。

関連するリンク
北海道大学和歌山研究林 クラウドファンディング Twitterアカウント
最新の活動報告
もっと見る
修論:引用文献一覧
2023/03/07 16:55引用文献一覧来週発送予定の修士論文の引用文献一覧です。皆様がアクセスしやすいように、この部分だけオンラインで共有させて頂きます。アルファベット順に並んでおります。本文は到着まで今しばらくお待ちいただければ幸いです。1. Agetsuma, N., Koda, R., Tsujino, R., & Agetsuma-Yanagihara, Y. (2016). Impact of anthropogenic disturbance on the density and activity pattern of deer evaluated with respect to spatial scale-dependency. Mammalian Biology, 81(2), 130–137. https://doi.org/10.1016/j.mambio.2015.09.0052. Anderson, J. T. (2009). Positive density dependence in seedlings of the neotropical tree species Garcinia macrophylla and Xylopia micans. Journal of Vegetation Science, 20(1), 27–36. https://doi.org/10.3170/2008-8-184883. Bagchi, R., Gallery, R. E., Gripenberg, S., Gurr, S. J., Narayan, L., Addis, C. E., Freckleton, R. P., & Lewis, O. T. (2014). Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature, 506(7486), 85–88. https://doi.org/10.1038/nature129114. Bardgett, R. D., Wardle, D. A., & Yeates, G. W. (1998). Linking above-ground and below-ground interactions: How plant responses to foliar herbivory influence soil organisms. Soil Biology and Biochemistry, 30(14), 1867–1878. https://doi.org/10.1016/S0038-0717(98)00069-85. Bayandala, Y. F., & Seiwa, K. (2016). Roles of pathogens on replacement of tree seedlings in heterogeneous light environments in a temperate forest: A reciprocal seed sowing experiment. Journal of Ecology, 104(3), 765–772. https://doi.org/10.1111/1365-2745.125526. Becklin, K. M., Pallo, M. L., & Galen, C. (2012). Willows indirectly reduce arbuscular mycorrhizal fungal colonization in understorey communities. Journal of Ecology, 100(2), 343–351. https://doi.org/10.1111/j.1365-2745.2011.01903.x7. Bennett, J. A., Maherali, H., Reinhart, K. O., Lekberg, Y., Hart, M. M., & Klironomos1, J. (2017). Population Dynamics Population Dynamics. In FOREST ECOLOGY (Vol. 69, Issue 2).8. Bever, J. D. (1994). Feeback between Plants and Their Soil Communities in an Old Field Community. Ecology, 75(7), 1965–1977.9. Bever, J. D., Westover, K. M., & Antonovics, J. (1997). Incorporating the Soil Community into Plant Population Dynamics : The Utility of the Feedback Approach Published by : British Ecological Society Stable URL : http://www.jstor.org/stable/2960528. Journal of Ecology, 85(5), 561–573.10. Canham, C. D. (1989). Different Respones to Gaps Among Shade-Tollerant Tree Species Author ( s ): Charles D . Canham Published by : Wiley on behalf of the Ecological Society of America Stable URL : https://www.jstor.org/stable/1940200 Ecological Society of America , Wiley are. EcologyEcology, 70(3), 548–550.11. Carteron, A., Beigas, M., Joly, S., Turner, B. L., & Laliberté, E. (2021). Temperate Forests Dominated by Arbuscular or Ectomycorrhizal Fungi Are Characterized by Strong Shifts from Saprotrophic to Mycorrhizal Fungi with Increasing Soil Depth. Microbial Ecology, 82(2), 377–390. https://doi.org/10.1007/s00248-020-01540-712. Carteron, A., Vellend, M., & Laliberté, E. (2022). Mycorrhizal dominance reduces local tree species diversity across US forests. Nature Ecology and Evolution, 6(4), 370–374. https://doi.org/10.1038/s41559-021-01634-613. Chabot, B. F., & Hicks, D. J. (1982). The ecology of leaf life spans. Annual Review of Ecology and Systematics. Volume 13, 229–259. https://doi.org/10.1146/annurev.es.13.110182.00130514. Chen, Y., Han, W., Tang, L., Tang, Z., & Fang, J. (2013). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography, 36(2), 178–184. https://doi.org/10.1111/j.1600-0587.2011.06833.x15. Cheng, X., Han, H., Kang, F., Liu, K., Song, Y., Zhou, B., & Li, Y. (2014). Short-term effects of thinning on soil respiration in a pine (Pinus tabulaeformis) plantation. Biology and Fertility of Soils, 50(2), 357–367. https://doi.org/10.1007/s00374-013-0852-016. Childs, S. W., & Flint, L. E. (1987). Effect of shadecards, shelterwoods, and clearcuts on temperature and moisture environments. Forest Ecology and Management, 18(3), 205–217. https://doi.org/10.1016/0378-1127(87)90161-717. Corrales, A., Mangan, S. A., Turner, B. L., & Dalling, J. W. (2016). An ectomycorrhizal nitrogen economy facilitates monodominance in a neotropical forest. Ecology Letters, 19(4), 383–392. https://doi.org/10.1111/ele.1257018. Crawford, K. M., Bauer, J. T., Comita, L. S., Eppinga, M. B., Johnson, D. J., Mangan, S. A., Queenborough, S. A., Strand, A. E., Suding, K. N., Umbanhowar, J., & Bever, J. D. (2019). When and where plant-soil feedback may promote plant coexistence: a meta-analysis. Ecology Letters, 22(8), 1274–1284. https://doi.org/10.1111/ele.1327819. de Jalón, L. G., Limousin, J. M., Richard, F., Gessler, A., Peter, M., Hättenschwiler, S., & Milcu, A. (2020). Microhabitat and ectomycorrhizal effects on the establishment, growth and survival of Quercus ilex L. Seedlings under drought. PLoS ONE, 15(6), 1–18. https://doi.org/10.1371/journal.pone.022980720. del Campo, A. D., Otsuki, K., Serengil, Y., Blanco, J. A., Yousefpour, R., & Wei, X. (2022). A global synthesis on the effects of thinning on hydrological processes: Implications for forest management. Forest Ecology and Management, 519(June), 120324. https://doi.org/10.1016/j.foreco.2022.12032421. Dickie, I. A., Davis, M., & Carswell, F. E. (2012). Quantification of mycorrhizal limitation in beech spread. New Zealand Journal of Ecology, 36(2), 210–215.22. Dickie, I. A., & Reich, P. B. (2005). Ectomycorrhizal fungal communities at forest edges. Journal of Ecology, 93(2), 244–255. https://doi.org/10.1111/j.1365-2745.2005.00977.x23. Ferriss, R. S. (1984). Effects of Microwave Oven Treatment on Microorganisms in Soil. In Phytopathology (Vol. 74, Issue 1, p. 121). https://doi.org/10.1094/phyto-74-12124. Gehring, C., & Bennett, A. (2009). Mycorrhizal fungal-plant-insect interactions: The importance of a community approach. Environmental Entomology, 38(1), 93–102. https://doi.org/10.1603/022.038.011125. Givnish, T. J. (2002). Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox. Silva Fennica, 36(3), 703–743. https://doi.org/10.14214/sf.53526. Hanberry, B. B. (2021). Addressing regional relationships between white-tailed deer densities and land classes. Ecology and Evolution, 11(19), 13570–13578. https://doi.org/10.1002/ece3.808427. Haskins, K. E., & Gehring, C. A. (2004). Interactions with juniper alter pinyon pine ectomycorrhizal fungal communities. Ecology, 85(10), 2687–2692. https://doi.org/10.1890/04-030628. Heinze, J., Wacker, A., & Kulmatiski, A. (2020). Plant–soil feedback effects altered by aboveground herbivory explain plant species abundance in the landscape. Ecology, 101(6), 1–10. https://doi.org/10.1002/ecy.302329. Hood, L. A., Swaine, M. D., & Mason, P. A. (2004). The influence of spatial patterns of damping-off disease and arbuscular mycorrhizal colonization on tree seedling establishment in Ghanaian tropical forest soil. Journal of Ecology, 92(5), 816–823. https://doi.org/10.1111/j.0022-0477.2004.00917.x30. Kadowaki, K., Yamamoto, S., Sato, H., Tanabe, A. S., Hidaka, A., & Toju, H. (2018). Mycorrhizal fungi mediate the direction and strength of plant–soil feedbacks differently between arbuscular mycorrhizal and ectomycorrhizal communities. Communications Biology, 1(1), 1–5. https://doi.org/10.1038/s42003-018-0201-931. Kayama, M. (2020). Growth and nutrient acclimation of evergreen oak seedlings infected with Boletus reticulatus in infertile colluvial soil in warm temperate Monsoon Asia: Evaluation of early growth. Forests, 11(8). https://doi.org/10.3390/f1108087032. Kayama, M., & Yamanaka, T. (2018). Growth of Quercus seedlings inoculated by ectomycorrhizal fungi at an abandoned site of reforestation. Journal of the Japanese Society of Revegetation Technology, 44(1), 33–38. https://doi.org/10.7211/jjsrt.44.3333. Kilpeläinen, J., Barbero-López, A., Vestberg, M., Heiskanen, J., & Lehto, T. (2017). Does severe soil drought have after-effects on arbuscular and ectomycorrhizal root colonisation and plant nutrition? Plant and Soil, 418(1–2), 377–386. https://doi.org/10.1007/s11104-017-3308-834. Kim, B. J., Oh, D. H., Chun, S. H., & Lee, S. D. (2011). Distribution, density, and habitat use of the Korean water deer (Hydropotes inermis argyropus) in Korea. Landscape and Ecological Engineering, 7(2), 291–297. https://doi.org/10.1007/s11355-010-0127-y35. Kliromonos, J. N. (2002). Feedback with soil biota contributes to plant rarity and invasiveness in communities. Nature, 417(May), 1096–1099. doi.org/10.1038/417067a36. Knoblochová, T., Kohout, P., Püschel, D., Doubková, P., Frouz, J., Cajthaml, T., Kukla, J., Vosátka, M., & Rydlová, J. (2017). Asymmetric response of root-associated fungal communities of an arbuscular mycorrhizal grass and an ectomycorrhizal tree to their coexistence in primary succession. Mycorrhiza, 27(8), 775–789. https://doi.org/10.1007/s00572-017-0792-x37. Kobe, R. K., Pacala, S. W., Silander, J. A., & Canham, C. D. (1995). Juvenile tree survivorship as a component of shade tolerance. Ecological Applications, 5(2), 517–532. https://doi.org/10.2307/194204038. Kulmatiski, A., Beard, K. H., Stevens, J. R., & Cobbold, S. M. (2008). Plant-soil feedbacks: A meta-analytical review. Ecology Letters, 11(9), 980–992. https://doi.org/10.1111/j.1461-0248.2008.01209.x39. Larson, J. L., & Siemann, E. (1998). Legumes may be symbiont-limited during old-field succession. American Midland Naturalist, 140(1), 90–95. https://doi.org/10.1674/0003-0031(1998)140[0090:LMBSLD]2.0.CO;240. Lee, E. H., Park, S. H., Eo, J. K., Ka, K. H., & Eom, A. H. (2018). Acaulosproa koreana, a new species of arbuscular mycorrhizal fungi (Glomeromycota) associated with roots of woody plants in Korea. Mycobiology, 46(4), 341–348. https://doi.org/10.1080/12298093.2018.154880541. Lodge, D. J. (1989). The influence of soil moisture and flooding on formation of VA-endo- and ectomycorrhizae in Populus and Salix. 117(2), 243–253.42. Ma, Y., Geng, Y., Huang, Y., Shi, Y., Niklaus, P. A., Schmid, B., & He, J. S. (2013). Effect of clear-cutting silviculture on soil respiration in a subtropical forest of China. Journal of Plant Ecology, 6(5), 335–348. https://doi.org/10.1093/jpe/rtt03843. Madritch, M. D., & Lindroth, R. L. (2011). Soil microbial communities adapt to genetic variation in leaf litter inputs. Oikos, 120(11), 1696–1704. https://doi.org/10.1111/j.1600-0706.2011.19195.x44. Maeda, M. (1954). The meaning of mycorrhiza in regard to systematic botany. 熊本大學理學部紀要. 第2部, 3, 57–84.45. Mangan, S. A., Schnitzer, S. A., Herre, E. A., MacK, K. M. L., Valencia, M. C., Sanchez, E. I., & Bever, J. D. (2010). Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature, 466(7307), 752–755. https://doi.org/10.1038/nature0927346. McCarthy-Neumann, S., & Iban, I. (2013). Reports Plant–soil feedback links negative distance dependence and light gradient partitioning during seedling establishment. The Ecological Society of America, 94(4), 780–786.47. Mcguire, K. L. (2007). Common Ectomycorrhizal Networks May Maintain. Ecology, 88(3), 567–574.48. McHugh, T. A., & Gehring, C. A. (2006). Below-ground interactions with arbuscular mycorrhizal shrubs decrease the performance of pinyon pine and the abundance of its ectomycorrhizas. New Phytologist, 171(1), 171–178. https://doi.org/10.1111/j.1469-8137.2006.01735.x49. Medve, R. J. (1984). the Mycorrhizae of Pioneer Species in Disturbed Ecosystems in Western Pennsylvania. American Journal of Botany, 71(6), 787–794. https://doi.org/10.1002/j.1537-2197.1984.tb14143.x50. Miyasaki, M., Sasaki, A., Kaneyuki, E., Ogura, A., Kinoshita, A., Nakatsubo, T., & Garden, B. (2015). 菌従属栄養植物ホンゴウソウの保全─生育環境の解明と移植─. Japanese Journal of Conservation Ecology, 220, 213–220.51. Mudrák, O., Hermová, M., Tesnerová, C., Rydlová, J., & Frouz, J. (2016). Above-ground and below-ground competition between the willow Salix caprea and its understorey. Journal of Vegetation Science, 27(1), 156–164. https://doi.org/10.1111/jvs.1233052. Murata, M., & Nara, K. (2017). Ectomycorrhizal fungal communities at different soil depths in a forest dominated by endangered pseudotsuga japonica. Nihon Ringakkai Shi/Journal of the Japanese Forestry Society, 99(5), 195–201. https://doi.org/10.4005/jjfs.99.19553. Nat Holland, J. (1995). Effects of above-ground herbivory on soil microbial biomass in conventional and no-tillage agroecosystems. Applied Soil Ecology, 2(4), 275–279. https://doi.org/10.1016/0929-1393(95)00055-254. Nishida, T., Izumi, N., Katayama, N., & Ohgushi, T. (2009). Short-term response of arbuscular mycorrhizal association to spider mite herbivory. Population Ecology, 51(2), 329–334. https://doi.org/10.1007/s10144-008-0116-255. Packer, A., & Clay, K. (2000). Soil pathogens and spatial patterns of seedling mortality in a temperate tree. Nature, 404(6775), 278–281. https://doi.org/10.1038/3500507256. Pangesti, N., Pineda, A., Pieterse, C. M. J., Dicke, M., & Loon, J. J. A. (2013). Two-way plant-mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Frontiers in Plant Science, 4(OCT), 1–11. https://doi.org/10.3389/fpls.2013.0041457. Putten, W. H. V. der, Dijk, C. van, & Peters, B. A. M. (1993). Plant-specific soil-borne diseases contribute to succession in foredune vegetation. Nature, 362(6415), 53–56. https://doi.org/10.1038/362053a058. Reich, P. B., Ellsworth, D. S., Walters, M. B., Vose, J. M., Gresham, C., Volin, J. C., & Bowman, W. D. (1999). Generality of leaf trait relationships: A test across six biomes. Ecology, 80(6), 1955–1969. https://doi.org/10.1890/0012-9658(1999)080[1955:GOLTRA]2.0.CO;259. Reynolds, H. L., Packer, A., Bever, J. D., & Clay, K. (2003). Grassroots ecology: Plant-microbe-soil interactions as drivers of plant community structure and dynamics. Ecology, 84(9), 2281–2291. https://doi.org/10.1890/02-029860. Seiwa, K., Negishi, Y., Eto, Y., Hishita, M., Masaka, K., Fukasawa, Y., Matsukura, K., & Suzuki, M. (2020). Successful seedling establishment of arbuscular mycorrhizal-compared to ectomycorrhizal-associated hardwoods in arbuscular cedar plantations. Forest Ecology and Management, 468(May), 118155. https://doi.org/10.1016/j.foreco.2020.11815561. Smith-Ramesh, L. M., & Reynolds, H. L. (2017). The next frontier of plant–soil feedback research: unraveling context dependence across biotic and abiotic gradients. Journal of Vegetation Science, 28(3), 484–494. https://doi.org/10.1111/jvs.1251962. Suding, K. N., Stanley Harpole, W., Fukami, T., Kulmatiski, A., Macdougall, A. S., Stein, C., & van der Putten, W. H. (2013). Consequences of plant-soil feedbacks in invasion. Journal of Ecology, 101(2), 298–308. https://doi.org/10.1111/1365-2745.1205763. Sveen, T. R., Netherway, T., Juhanson, J., Oja, J., Borgström, P., Viketoft, M., Strengbom, J., Bommarco, R., Clemmensen, K., Hallin, S., & Bahram, M. (2021). Plant-microbe interactions in response to grassland herbivory and nitrogen eutrophication. Soil Biology and Biochemistry, 156(October 2020). https://doi.org/10.1016/j.soilbio.2021.10820864. Swaty, R. L., Deckert, R. J., Whitham, T. G., & Gehring, C. A. (2004). Ectomycorrhizal abundance and community composition shifts with drought: Predictions from tree rings. Ecology, 85(4), 1072–1084. https://doi.org/10.1890/03-022465. Takashima, T., Hikosaka, K., & Hirose, T. (2004). Photosynthesis or persistence: Nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant, Cell and Environment, 27(8), 1047–1054. https://doi.org/10.1111/j.1365-3040.2004.01209.x66. Tedersoo, L., Bahram, M., & Zobel, M. (2020). How mycorrhizal associations drive plant population and community biology. Science, 367(6480). https://doi.org/10.1126/science.aba122367. Trocha, L. K., Weiser, E., & Robakowski, P. (2016). Interactive effects of juvenile defoliation, light conditions, and interspecific competition on growth and ectomycorrhizal colonization of Fagus sylvatica and Pinus sylvestris seedlings. Mycorrhiza, 26(1), 47–56. https://doi.org/10.1007/s00572-015-0645-468. van der Putten, W. H., Bardgett, R. D., Bever, J. D., Bezemer, T. M., Casper, B. B., Fukami, T., Kardol, P., Klironomos, J. N., Kulmatiski, A., Schweitzer, J. A., Suding, K. N., van de Voorde, T. F. J., & Wardle, D. A. (2013). Plant-soil feedbacks: The past, the present and future challenges. Journal of Ecology, 101(2), 265–276. https://doi.org/10.1111/1365-2745.1205469. Wang, B., & Qiu, Y. L. (2006). Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza, 16(5), 299–363. https://doi.org/10.1007/s00572-005-0033-670. Wang, Y., Li, Z., Wang, S., Wang, W., Wang, N., & Gu, J. (2022). Variations in Arbuscular Mycorrhizal Colonization Associated with Root Diameter and Hypodermis Passages Cells across Temperate and Tropical Woody Species. Forests, 13(2), 8–11. https://doi.org/10.3390/f1302014071. Wright, I. J., Westoby, M., Baruch, Z., Bongers, F., Cavender-Bares, J., Chapin, T., Diemer, M., Others, Wright, I. J., Reich, P. B., Ackerly, D. D., & Cornelissen, J. H. C. (2004). The worldwide leaf economics spectrum. Nature, 428, 821–827. www.nature.com/nature%0Ahttps://idp.nature.com/authorize/casa?redirect_uri=https://www.nature.com/articles/nature02403&casa_token=CtYdn6u63BcAAAAA:cNAkoHi669d547DoKaM3keUixX15dhKLnmyUD_oX2GBEB1VTe9mtWcMdFoXauJPFpb4iQItw5D0oAvb372. Wulantuya, Masaka, K., Bayandala, Fukasawa, Y., Matsukura, K., & Seiwa, K. (2020). Gap creation alters the mode of conspecific distance-dependent seedling establishment via changes in the relative influence of pathogens and mycorrhizae. Oecologia, 192(2), 449–462. https://doi.org/10.1007/s00442-020-04596-x73. Yang, A. N., Lu, L., & Zhang, N. (2011). The diversity of arbuscular mycorrhizal fungi in the subtropical forest of Huangshan (Yellow Mountain), East-Central China. World Journal of Microbiology and Biotechnology, 27(10), 2351–2358. https://doi.org/10.1007/s11274-011-0702-x74. 吉岡広美, 磯部香, 乾久子, & 菊地淳一. (2012). アベマキ-アラカシ混交林の外生菌根菌相と子実体の空間分布. 奈良教育大学自然環境教育センター紀要, 13, 1–13. http://hdl.handle.net/10105/923475. 林野庁. (2021). 平成14年3月29日 13林整整第882号 農林水産事務次官依命通知 最終改正:令和3年3年31日 2林整整第946号 第1 趣 旨.76. 櫻庭知帆. (2018). 哺乳類分布の季節変動を規定する要因は何か-狭い範囲で 環境が大きく異なる生息地において -. https://www.ptonline.com/articles/how-to-get-better-mfi-results もっと見る
12月後半活動報告
2022/12/31 19:17こんにちは!先日、西日本の大雪がニュースになっていましたが、古座川でも一部の地域で雪が積もりました。かなり大粒の雪が結構な勢いで降り続いていたので、北海道のような光景でした。古座川の七川ダムより上流の地域は、町の中では標高が高く、気温も下がりやすい地域なので、雪が積もるとしたらダムより上流というイメージがありました。ところが今回は、ダムより下流域で積もったので珍しい光景を見ることができました。定期報告 最終回さてさて、今回の活動報告は定期報告としては、いよいよ最終回となります。 僕は年末で平井を離れ苫小牧に行くので、平井での生活を報告できるのも今回が最後になります。そこで、研究のまとめは年度末に行うことにして、今回は平井での生活を振り返ってみたいと思います。春平井と言っておきながら、最初から別の集落の写真です。都市部でサクラと言えば並木にされて群れているソメイヨシノですが、古座川では点在するようにヤマザクラやクマノザクラが分布しています。サクラは、密集して生えていると、分かりやすい華やかさがありますが、ポツンと生えている様子も、幽玄な雰囲気を醸していて綺麗でした。色もやや薄めのピンクですが、ほんのりとした色合いは、照葉樹の濃い緑に良く映えていました。こちらは、近所にあった和歌山県最古の木造校舎です。つい最近解体されてしまいましたが、長年連れ添った校庭の桜だけは今も残っています。茶摘みゴールデンウィークに入ると、茶摘みでした。平井では、庭の生垣にお茶が植えられているので、集落の方々は、その新芽をもぎ取ってお茶にしています。一番茶を採った後は、本格的な生産地なら二番茶、三番茶と続きます。しかし、平井では一番茶だけで事足りるので、「贅沢やぁ~」と集落の方も仰っていました。取ってきたお茶っ葉は、大きな鉄鍋で煎ります。このお婆ちゃん曰く、「鉄じゃなきゃ美味しくならんよ」とのことでした。そのため、一家に一台大きな鉄釜を買って、錆びないように大事に屋根の下で保管するのだそうです。大きな釜にいっぱいの茶葉を入れて、ゆっくりと煎っていきます。かなり熱そうですが、手慣れた手つきで淡々と作業をするばあさん。この光景を知っていると、尚更お茶が美味しくなりそうです。実際にできたお茶がこちら。非売品ですが、研究林に来れば飲めるかも?田植えお茶摘みが終わったら田植えが始まります。新潟平野の広大な田んぼのようなものはなく、小さな棚田が少ない平地に何枚かある程度です。こんなに尾根の近いところで水田が広がっているのは、平野で育った身としては新鮮な光景です。尾根に近いのに、田んぼに水を張れるだけの量の水をどこから引いているのか、そんな疑問が湧いてきます。平井の場合は、裏手の山から水を引いているわけではなく、川の上流で標高の高いところから、なるべく平行に用水路を掘って水を引いています。原理的には玉川上水と一緒です。棚田の間には、石段の路地が張り巡らされており、箱庭のようになっています。そこに北大の青い屋根が遠くに見えると、何ともメルヘンチックでした。田んぼにはアカハライモリやカエルが遊びに来ています。田んぼがあるからこその生態系が築かれており、人々の生活と周辺環境との長い間続いた相互作用を想起させます。夏の平井梅雨頃になると、ホタルが小川を乱舞します。ここほど大量にホタルが飛んでいるのは、ほかでは見たことありません。乱舞という言葉も古座川のホタルを見て、やっとしっくりきました。2022年の夏は、十数年ぶりに平井の夏祭りが開かれました。捻りが特徴の平井伝統の盆踊りも復活して、お囃子が山あいにこだましていました。来年も開催されたら、是非参加したいですね。普段の夏はこんな感じ。雨でしっとりと濡れたい地面や、低い雲が熊野の深い山の雰囲気を作り出します。お盆では、送り火として非常に高い竹の先に松明を付けて灯します。他の地域では見たことが無かったので、盛大な送り火には驚きました。送り火の頃は、雨も多いので、一瞬止んだ隙を狙って立てていました。背景の雲がかかった山々も相まって、信仰の土地であることを再認識させられる雰囲気です。翌日には、精霊流しがありました。河原で線香を焚いてから、ご先祖さまが帰るときのお弁当を流します。平井の川は激流なので、お弁当もとんでもない勢いで流れていったのが衝撃的でした。お盆もすぎると、秋になります。秋はオオサンショウウオの繁殖期で、河原をオオサンショウウオがペチペチ歩いていました。初めて見たオオサンショウウオ。ぷにぷにの手足がめちゃくちゃキュートでした。稲刈同じころ、夏前に植えた稲の刈り取りが始まります。ちょうど台風がたくさんやってくる時期なので、この作業も天気との勝負になります。2022年に平井で一番広い田んぼに植えられた稲の品種は、早く育つ種だったようです。そのため、例年よりも色付きが早く、まだ暑いなか収穫が始まりました。収穫は、田んぼの持ち主だけでなく、親族や近所の方々が手伝いに来てくれます。区画が直線的でなく、田んぼの面積も小さいので、大型機械には不向きです。そのため、こうした方々の協力は欠かせません。普通に買った方が安いかもしれないけど、楽しいから続けているという家もあるようです。柚子とり稲刈が終われば、最後は柚子とりです。柚子の里を名乗る平井は、そこらじゅうに柚子が植わっており、秋も終わりになると黄色い実が目立ってきます。収穫された柚子は、平井の集落内にある工場で絞られます。そのときに、柚子の香りが集落中に広がり、季節を感じさせる風物詩になっていました。2022年は、僕も収穫に参加させて頂きました!柚子の木の下でおやつを食べる時間は、色々な集落のお話を聞くことができ、非常に楽しい時間でした。特に印象的だったのは、上の写真の方が教えてくれた、サルの脳みその焼き方でした笑。なんでも、赤土で覆ってから鍋に入れてじっくりと焼くそうです。頭痛の薬に使っていたんだとか。冬柚子の収穫が終われば、もう冬です。住むまで知りませんでしたが、本州最南端と言えど山間に位置する平井は、意外に雪が降ります。特に2021年から2022年にかけての冬は、5回ぐらい積もったうえに、-6℃ぐらいまで下がった日もあったので、札幌とあまり変わりませんでした。雪の光景と、北大チックな庁舎はやっぱり似合いますね。そんなこんなで年を越えると、ぼちぼち梅が咲いてきます。梅が咲けば、また新しい1年がやってきます。このような1年が、この平井で年十年、何百年と繰り返されてきたのだなぁと思うと、その歴史の一部に参加することができてよかったと感じます。他の山村と同じように、平井もやはり深刻な高齢化に悩まされている地域です。今後20~30年で、日本中の似たような地域のほとんどが消滅してしまうことを考えると、ここで僕が見て感じたことは、50年後60年後には非常に貴重な体験談となることでしょう。そのときも、平井の地に人が住み続けていることを願いながら、定期活動報告を締めたいと思います。なお、研究に関する報告書は年度末(3月ごろ)に発送いたします。そちらでも、平井の様子をまとめたものを作る予定ですので、どうぞ楽しみにお待ちください。それでは、非常に長い間、活動報告を読んでいただき有難うございました!機会があれば、是非古座川町の平井に遊びに来てください!2022年12月31日北海道大学和歌山研究林井口光 もっと見る
12月前半活動報告
2022/12/15 21:55こんにちは。寒さが身に染みるようになり、お風呂が楽しみな季節になってきましたね。昨日はちょうど流星群が見れたようで、いつもより多くの流れ星が飛び交っていました。こんな星空も、あとちょっとと考えると名残惜しい気持ちでいっぱいです。平井の生活も残すところあと2週間なので、ぼちぼち荷物をまとめています。さて、今日は、前回の続きのアートイベントからです。森のちからⅩⅢ 森の聲前回は古座川町に在住の南条さんの作品を紹介しました。今回は、同じくゲストで来て頂いた彫刻家の吉野祥太郎さんの作品を紹介します。 一つ目の作品は、研究林に入って一つ目の物奥小屋にありました。実はまだ1回も入ったことがなかったので、どんな展示になっているのか楽しみです。中に入ってみると、枯山水を思わせるような空間になっていました。枯山水と違うのは、本物の水が沢から引かれて、水の滴る音が響いている点です。澄んだ空気と相まって、心穏やかになりそうな雰囲気でした。彫刻家と聞いていたので、てっきり何か彫った作品が置かれているのかと思っていましたが、インスタレーション的に幅広く制作されているようです。作品のタイトルは「森の聲」。研究林周辺の道路に落ちてきた岩や、動物たちが蹴り落とした小石、降ってきた枯れ枝などが、単なる偶然ではなく彼らの意思でそこに来たとしたら?というコンセプトだそうです。そのため、水に浮かんでいる木や岩は、道に落ちていたものをそのまま持ってきたものなんだとか。僕の研究も、一見確率論的に見える実生の分布に隠された森のルール(意思?)を探るようなものなので、何か共通する部分を感じます。以前、尾根のてっぺんの方に、重い種子を持つトチノキの実生がポツンと生えていたのをご紹介しましたが、あの個体を発見した時は、「なんでここにいるの?」と聞きたくなりました。そこに”在る”ようになるまでの様々なストーリーを考えると、いろんな想像が膨らみます。それは、落石一つとっても同じことかもしれません。それにしても、建物の中に水が流れ野性味あふれる落石や伐根が転がっているのは、なかなか面白い光景です。特に水の流れ。子どものころ、水が流れているのを見るのが大好きで、よく湧き水を見に行ったり、庭で水遊びしていたりしましたが、24になっても相変わらず水の流れには心ときめきます!普通は外でしか見ることができない流水を、屋内で見られるというそれだけで僕は大興奮でした!この興奮は人間の住環境が、屋内と屋外で明確に区切られた文化が形成されたからこその、驚きともいえるかもしれません。ただ、人間が自然を完全に排除した空間に耐えられなくなり、盆栽や観葉植物のように、自然を持ち込もうとする行為が許されるのであれば、自然側の意思を持って人間界へ浸食することも否定できないかもしれません。そのような自然の意思による浸食が、落石や落枝落葉となって、今日も道に積もっているのかもしれないなぁ、なんて思いました。水による記憶時計装置次の作品は橋を渡った先の生け簀跡です。このロケーションが既に作品の一部のようになっていますね。今日も川は綺麗です!中はこのようになっていました。こちらの作品は、写真だとなかなか伝わりにくくて申し訳ないのですが、中央に落ちている水が、点滅する光によって、ときに規則的に、ときに不規則に照らされていました。解説によると、人間がろ過するのとは異なる、「山による壮大な濾過を経た水」がコンセプトのようです。それを聞いてから改めて作品を見てみると、なるほど、この暗い空間は森林の下で繰り広げられている、水の旅路なのかと腑に落ちました。ともすれば天井から垂れ下がっている植物の一部も、木々の根っこのように解釈することもできます。雨として降った水は、多くが木々に吸い上げられ、蒸散によって空気中へ戻っていきます。そうした水とは異なるルートを選んだ水たちは、ゆっくりゆっくりと深いところへ沈んでゆき、森の深いところで涵養されています。そうした水が辿ってきた旅路の記憶を具現化したような作品だと感じましました。この作品を見て思い出したのは、「真珠のボタン」と呼ばれるチリのドキュメンタリー映画です。作品自体は、チリ沖の海底で見つかった真珠のボタンから、チリの民族差別や独裁政治など負の歴史を顧みる構成になっています。興味深いのは、タイトルこそ「真珠のボタン」となっていますが、作品全体は「水の記憶」で括られていることです。作品の冒頭、「水は宇宙からやってきて、生命の誕生から現在に至るまで、全てを見てきた…」と始まり、最終的に真珠のボタンが沈んでいた海、つまりは水は人々の蛮行をずっと見ていた、と終わります。真珠と水をパラレルな関係におくことで、真珠の記憶すら凌駕する、水のストーリーの壮大さが際立っています。「物が記憶する」というのは一聞では、「まさか」と思いますが、木の年輪や地層、南極氷河に含まれる気泡は、気の遠くなるほどの歴史を確かに刻んでいます。それと比べたら人間の記憶なんて些細なものですから、記憶が人間の特権なんて考えは驕りでしかないかもしれませんね。小学生たちの作品作品作りをしているのは大人だけではありません。川下の小学校で木育活動をしていると、結構な頻度で「おもしろい!!」と思うような作品を作る方がたくさんいます。今回は、この1年の木工教室で特に面白かった作品をご紹介しようと思います。例によって、材料は北大産の材です。不要になった材の山から、使えそうなのを引っ張ってきて、輪切りにしたり板にしたりして再利用しています。輪切りを作るのが本当に大変で、何十枚と切るので、細い枝材と言えど筋肉痛になりました。まずはこちら。絶妙なバランス感覚が光っています。板は床か壁に使うものだという固定観念を覆し、柱として利用しているのもポイントが高いですね。多様な樹種が取り入れられているのも、現代的な問題意識が現れていて素晴らしい作品です。ちなみに、これを作った子は持って帰るのを忘れてしまい、僕が引き取りました。お次はこちら。この作品を作った子は、いっつも面白いキャラクターを作ってくれます。今回の作品も、異形のものかと思いきや、どこか可愛らしい際どいラインを見事に攻めた作品を見せてくれました。ちなみに、この子は作った作品をいつも僕にくれるので、我が家へ持って帰りました。続いてこちら。三日月のような部分はヤマグワの樹皮でできています。なかなか面白い材を見つけたものだと感じました。 月面の静寂を感じさせる洗練された配置です。近未来的でSFチックな世界観を、木という人工的な素材とは真逆の素材によって完成させたことに、驚きを感じます。人為とは何なのか?を問いかけるかなり示唆深い作品です。こちらは家のモデルですね。脱炭素化の加速のためには木造化が欠かせないというメッセージを感じます。二股に分かれた大きなツバキの材が、単調になりそうな家の雰囲気をガラリと変えています。自然への畏敬の念を忘れるなということでしょうか。こちらはシカを大胆に解釈し、再構築した作品ですね。枝の切跡に上手く目を付けた秀逸な作品です。シカの目が側面を向いていることも理解して作られていて、小学生にして優れた観察眼を持っていることが分かります。学校変わってこちらはペン立てだそうです。飛び込み台のように見えるのは消しゴムを置くスペースとのこと。「僕には絶対そんな発想ができない」と思わされました。側面に張り付いているのは、クロモジとヤブニッケイの輪切りです。どちらもクスノキ科でよい香りがします。こちらもペン立て。配色にセンスが光っていますね。明るい部分には明るい色で飾りをつけ、舌の暗い樹皮にはクロモジと濃い色で装飾を施しており、適当ではない作為が感じられます。こちらではどんぐりとナンテンを使っての工作です。一番面白かったのは、キャラクター職人の彼。今回は、どんぐりの帽子をナンテンの実に取り付け、枝に付け直すという離れ業をやってくれました。どんぐりを使うか、ナンテンを使うかで悩むのではなく、その融合を達成する心意気。ヘーゲルもびっくりです。こちらはシンプルですが、彼しかやっていなかった「どんぐり二段重ね」が目を引きました。まさか、どんぐりが縦に重なるなんて思わないじゃないですか!重なってるどんぐりもびっくりしてることでしょう。森林圏交流会12月前半は、僕の所属する北大環境科学院の森林圏科学コースで交流会があり、中間報告を兼ねて研究発表を行ってきました。まだ、一般の方向けの解説を書けていないのですが、参考までに当日使用したスライドと、原稿をご紹介したいと思います。暫定的に「優占種の菌根菌タイプは定着する実生の動態に影響を及ぼすか?」というタイトルで発表しました。種の共存メカニズムの解明は、生態学の重要なテーマです。樹木では、成木が実生に及ぼす密度依存的な効果が共存に重要だと考えられており、この密度依存性は植物と土壌のフィードバックに影響を受けています。植物ー土壌間のフィードバックとは、成木が土壌に対し生物的/非生物的に作用し、形成された土壌が周囲の植物個体のパフォーマンスに影響することを指します。この相互作用を、以降、PSFと呼びます。成木と実生の種の一致/不一致でPSFの質が異なるとき、PSFは森林動態や群集構造に影響を及ぼします。例えば、共生菌の蓄積は、同種の定着を促進し、優占種の優位性を高めて多様性を低下させる可能性があります一方、成木の近くに同種に特異的に感染する土壌病原菌が蓄積されると、優占種の定着が減少し、群集の多様性を高めることができます。このPSFの初期の研究は、実生の樹種が、土壌を形成した樹種と一致するかどうかで、PSFの性質が異なるか種ごとに検証するものでした。しかし、最近の研究では、実生の菌根菌のタイプによって、PSFの性質を説明できることが示唆されています。例えばBennettらは実生の共生菌が、外生菌根菌(AM)の場合は正,アーバスキュラー菌根菌(EM)では負のPSFが生じることを実験で明らかにしました。またKadowakiらは、樹種の同一性と、実生の菌根型に加えて、土壌を形成した樹種と実生の、菌根型の同一性でもPSFがあることを示しました。これによって、例えばAM型は同種では負のPSFがあっても、菌根型では正のPSFが生じていることが示されました。菌根型の同一性でPSFが生じるのであれば,群集における優占種の菌根型は定着する実生の動態に群集規模で影響を及ぼしているのでしょうか?例えば和歌山研究林の人工林はアーバスキュラー菌根菌と共生するスギ・ヒノキが優占種。天然林は外生菌根菌と共生するコナラ属が優占種となっています。Kadowakiら(2018)で示されたPSFが生じていれば、AM型樹木の実生は人工林内で正、天然林内で負のPSFが生じており、EM型は逆のパターンになると考えられます。しかし, フィールドでは昆虫や哺乳類などが、密度依存的な負の効果をもたらすことが報告されています。そのため、種子散布数の多い天然林は実生に負の影響が生じやすいと考えられ、実生の動態に影響を及ぼす要因を分離して検討する必要があります。そこで、本研究ではまず初めに①フィールドの土壌は優占種の菌根型によって条件付けされているのか?を温室での殺菌処理実験により検証し、次に②フィールドの実生の動態が菌根型によるPSFで説明可能であるか、をフィールドにおける毎木調査で検証しました。ただし、地上部の昆虫の被食圧は、菌根菌への依存度を高め、種レベルのPSFを強めることが報告されています。PSFが地上部の食害に起因する場合、土壌処理だけではPSFを検証できない可能性があります。そこで、温室実験では、土壌処理と共に模擬食害を行うことで、地上部の食害がPSFの発生要因になっているか検討しました。本研究の仮説です。1つ目が菌根菌を介したPSFは、地上部の食害に起因する2つ目がフィールドの土壌は優占種の菌根型によって条件付けされている具体的には、AM型実生においては天然林よりも人工林でPSFが正になり、EM型実生は負になる。という仮説を立てました。方法です。調査地は北大和歌山研究林で、人工林と天然林が隣接する場所を4ヵ所選定しました。人工林はスギ・ヒノキ、天然林はコナラ属の優先する照葉樹林です。まず、この4地点で土壌採取を行いました。土壌を採取したのは林縁から人工林・天然林へそれぞれ30mの地点と、中間地点です。採取した土壌を篩にかけ、ばらつきを減らすため4ヵ所の土をよく混合しました。混合した土壌は半分を電子レンジで殺菌しました。次に、EM型の実生として、アカガシを、AM型の実生としてヤマザクラを、それぞれ表面殺菌して、ポットに植えました。植えた実生の半分に対し、葉面積の8%に穴が開くよう、模擬食害処理を行いました。測定項目は樹高と葉面積で、実験開始前と、10週間経過後の2回測定を行いました。まとめると、処理は大きく3つあります。一つ目が土壌の採取地点で、天然林・人工林・中間地点です。2つ目が殺菌処理で、3つ目が模擬食害処理です。これらを掛け合わせた計12種類の苗に対し、各20反復ずつポットを作り温室でランダムに配置しました。解析方法です。PSFの方向性と強さを処理別で比較するため、二元配置分散分析をしました。応答変数はPSFの指標として、殺菌していないポットでの成長量を、殺菌したポットでの成長量で割り、対数をとった値としました。説明変数は、土壌を取った場所の森林タイプと模擬食害に加え交互作用も考慮しました。森林タイプもしくは交互作用に有意差が見られた場合はTukeyの多重比較を行いました。まずAM型のヤマザクラです。仮説では、PSFは食害に起因しAM型は天然林よりも人工林でPSFが正になるとしました。グラフは、横軸が森林タイプで、縦軸がPSFの指標となっています。縦軸は0より上でPSFが正、0より下でPSFが負になることを意味します。箱ひげの色は黄色が食害アリ、オレンジが食害ナシの結果となっています。二元配置分散分析を行った結果、食害処理には有意差が見られず、森林タイプに有意差があり、天然林が人工林に対し、有意に低いことが分かりました。次にEM型のアカガシの結果です。仮説では、天然林で人工林よりもPSFが正になるとしました。結果は、食害処理では有意差が見られず、森林タイプでのみ有意差が見られ、人工林が天然林よりも有意に低いことが分かりました。まとめと考察です。PSFの発生原因だと考えていた食害は、PSFに対し有意な影響を及ぼしていませんでした。このことから、PSFは食害の有無に関係なく生じていると考えられます。次に、森林タイプの効果です。AM型では人工林よりも天然林で負の影響EM型では正の影響がありました。この傾向は菌根菌のPSFから予想されるパターンと一致しているため、フィールドの土壌は優占種の菌根型によって条件付けされていることを支持する結果となりました。続いて、フィールド調査についてです。フィールド調査では、温室で見られた菌根型によるPSFの違いが、フィールドにも当てはまるのか検証します。実生の動態が菌根型によるPSFで説明可能であるならばAM型のパフォーマンスは人工林で良好で、EM型は天然林で良好であると予想できます。温室と異なり、フィールドで異質性の見られる光環境や水分環境などは、PSFに影響を及ぼすことが分かっています。特に日本では近年、種多様性保全のために人工林で間伐が行われています。間伐は人為的撹乱として、光環境や土壌含水率を変化させることから、PSFに影響を与えている可能性が高いと考えられますしかし、間伐が菌根型によるPSFに及ぼす影響を検証した研究はほとんどありません。より正確に森林動態を予測するためにもPSFの側面から間伐を検討する必要があります。例えば間伐による光環境の改善は、菌根菌のコロニー形成率を上昇させたり、AM型実生の病原菌耐性を向上させたりします。しかし、(ボタン)間伐強度が高くなってくると、土壌水分が減少し乾燥ストレスがかかります。乾燥ストレスに対する菌根菌の応答は、AMとEMで異なっていることが報告されており、EMはコロニー形成率を低下させるのに対し、AMはEMよりも乾燥耐性が強く、乾燥条件下で共生する利益が最大化することが報告されています。したがって、間伐がPSFに与える影響は菌根型によって異なることが考えられます。以上のことから、フィールド調査においては、大きく2つの仮説を立てました。1つ目は、フィールドにおける実生のパフォーマンスです。AM型は天然林よりも人工林で良好、EM型では逆という仮説を立てました。二つ目は、間伐が、菌根型によるPSFに及ぼす影響についてです。AM型は間伐強度の増加に伴い、人工林での正の効果が向上する。EM型は天然林での正の効果が低下すると仮説を立てました。方法です。調査地は土壌採取地点と同じです。間伐は列状間伐とし、強度を4つ、コントロール、5m幅, 10m幅, 20m幅を各4rep.設置しました。間伐区の長さは40mで、バッファーは15mとしました。毎木調査は林縁から人工林と天然林にそれぞれ30m進んだ地点と、人工林に5m進んだ、計3地点で行いました。プロットは地図上で赤い四角で示しています。毎木調査は2年間で計4回行いました。低木・灌木・つる性・ツツジ科木本を除く50㎝未満の木本植物を対象として、樹高と、生存状況を測定しました。調査対象になった樹種は全部で46種です。これらを先行研究を基に、表のようにAM型・EM型に分類しました。解析に使用したデータについて補足します。フィールドにおける菌根型によるPSFの検証では、天然林と無間伐人工林の30m地点のデータを使用しました。黄色で示したプロットです。一方、間伐がPSFに及ぼす影響の検証では、天然林での伐採が許可されていないため、人工林内のデータを用い、天然林からの距離に対する実生の応答が、菌根型によって異なるか検証しました。使用したデータはピンクの〇で囲った範囲のものです。結果です。まずはフィールドおける菌根型によるPSFの検証です。解析はGLMMで行い応答変数を、2年間の生存率。説明変数を、初期樹高と森林タイプとしました。AICが最小のものをベストモデルとしました。結果は、AM型においては、森林タイプが選択され、天然林で負の効果が見られたのに対し、EM型では、森林タイプがベストモデルに選択されませんでした。同じように、樹高の成長量についてGLMMで解析したところ、成長量では、AM型・EM型ともにベストモデルで森林タイプが選択されませんでした。考察です。AM型では生存率が人工林の方が良く、菌根型によるPSFの仮説を支持しました。一方、EM型では傾向が見られず、菌根型によるPSFで実生の動態を説明できませんでした。AM型・EM型ともに成長量で傾向が見られなかったのは、調査期間が短く、充分な成長量が測定できなかった可能性が考えられます。また、EM型で傾向が見られなかったのは、EM型でパフォーマンスが良いと予想していた天然林において、シカ食害がより強い制限要因になっている可能性が考えられます。実際、シカは針葉樹人工林を忌避することが報告されており、和歌山研究林でも同様の傾向が報告されています。そのため、EM型の天然林における優位性が,シカ食害により相殺され森林タイプに差が生じなかった可能性が考えられます。次に、間伐が菌根型によるPSFに及ぼす影響についてです。GLMMの解析の説明変数に、間伐強度と天然林からの距離をおき、交互作用項を含めて、先ほどと同様に解析を行いました。まず生存率についての結果です。AM型の生存率では、間伐強度や天然林からの距離がベストモデルに選択されませんでした。一方、EM型では、間伐強度で負の影響、天然林に近い場所で正の影響が見られ、交互作用項が選択されました。EM型の交互作用項について、平均限界効果を求めたところ、間伐強度が20mの地点で、天然林付近での優位性が低下していることが分かりました。続いて成長量です。先ほどまでと同様に解析を行いました。その結果、AM型においては間伐が正の影響、天然林の近くが負の影響を及ぼしていることが分かりました。一方EM型においては、間伐が選択されず、天然林の近くで正の影響があることが分かりました。考察です。天然林からの距離の効果は、AM型では生存率には影響しなかったものの、成長量では正の影響がありました。一方、EM型は天然林からの距離が生存率・成長量に負の影響を与えていました。この傾向は、概ね温室実験と合致し、菌根菌PSFの仮説を支持する結果となりました。天然林で行った調査とは異なる結果が生じたのは、シカの食害を受けない範囲だったためだと考えられます。また、間伐は、AM型の成長量を増加させましたが、天然林からの距離の効果には影響を及ぼしていませんでした。一方EM型では、間伐によって天然林の近くでの優位性が低下していました。先行研究ではAMの方がEMよりも乾燥耐性があることが示されています。そのため、AM型では強度間伐による乾燥に耐えることができたものの、EM型では、乾燥で共生する菌根菌が減少し、PSFのベネフィットが減少したと考えられます。まとめです。フィールドの土壌は優占種の菌根菌によって条件付けされていました。この結果は菌根菌によるPSFの存在を支持しています。しかし、フィールドにおいては人工林内でのみ菌根型によるPSFを支持する傾向が見られましたことから、フィールドでの影響力は局所的であると考えられます。また、間伐は、菌根型によって天然林からの距離の効果に異なる影響を与えていました。AM型では影響が見られなかったのに対し、EM型では天然林付近の正の効果が間伐強度の増加に伴い低下していました。ということで、中間発表が終わったので最終発表と修論執筆に本腰を入れていきたいと思います。次回が定期報告は最終回となりますが、どうぞお楽しみに! もっと見る






コメント
もっと見る